
Tato studie zkoumá vliv hedvábného vlákna na lidský organismus, popisuje jeho příznivé vlastnosti a také jeho účinnost v boji proti některým nemocem, díky jeho chemické struktuře. Tato práce popisuje a analyzuje možnosti zlepšení hedvábí a také výhody jeho dalšího používání. Studie obsahuje podrobný přehled řady vědeckých prací, z nichž vychází.
Hedvábí je souvislé proteinové vlákno produkované bourcem morušovým k uzavření jeho zámotku (Cook, 1968). Někteří pavouci z čeledi Arachnida jsou také schopni produkovat hedvábné vlákno (Cook, 1968; Robson, 1985). Jejich vlákno však nepodléhá průmyslové výrobě. Proto se tato studie zaměří na hedvábí vytvořené výhradně bourcem morušovým (bourem morušovým).

Na rozdíl od vlny obsahuje hedvábí velmi malé množství síry (Dhavalikar, 1962). Existují dvě klíčové odrůdy hedvábí: morušové hedvábí (vyrábí Bombyx Mori), také nazývané „kultivované“ hedvábí, a „divoké hedvábí“, jehož nejběžnější odrůdou je Tussah.
Morušové hedvábí produkují bource morušového chované ve speciálně určené oblasti a krmené čerstvě natrhanými listy moruše. Vlákno pěstovaného hedvábí je velmi tenké, téměř bílé (pokud není slepeno) s malými skvrnami jemného lesku.
Divoké hedvábí Od moruše se liší především tím, že divoký bourec morušový se živí hlavně dubovými listy. Je hrubší, nerovnoměrnější a má nahnědlý nebo žlutý odstín (Sandoz Color Chronicle, 1990).
Téměř 80–85 % světové produkce hedvábí pochází z morušového hedvábí.
Schopnost bource morušového produkovat 1 až 8 generací (sklizně kokonů) za rok je jednou z vlastností, kterými se plemena bource morušového liší. Existují tři příbuzné druhy bource morušového (Venugapal, 1991).
Monovoltinské plemeno distribuován především v Evropě. Vzhledem k relativně chladnému klimatu jsou vajíčka snesená samicí přes zimu ve stavu diapauzy neboli „hibernace“ a k oživení a oplodnění dochází až na jaře. Proto bourec morušový produkuje pouze jednu generaci za rok.
Bivoltine plemeno typické pro země jako Japonsko, Čína a Korea. Tato odrůda se od předchozí liší tím, že díky teplejšímu klimatu je schopna produkovat dvě generace ročně. Motýli bivoltinních plemen jsou menší než univoltinní, kvalita zámotků svinutých housenkami je nižší, ale bivoltinní plemena jsou předčasná a odolná. Křížením bivoltinních plemen s monovoltinními plemeny, která produkují hedvábí nejvyšší kvality, byla získána nová plemena bource morušového, vyznačující se vysokou životaschopností, velkými rozměry kokonů, vysokou produkcí hedvábí a dobrými technologickými kvalitami hedvábí (Michajlov, 1956; Kovalev, 1956) .
Multivoltinové plemeno vyskytují výhradně v tropech. Jakmile samička bource morušového naklade vajíčka, jejich vývoj trvá 9 až 12 dní, což má za následek 8 samostatných životních cyklů za rok. Většina bource morušového patří k mono- a bivoltinním plemenům.

STRUKTURA HEDVÁBNÉHO VLÁKNA
Předení bource morušového popsali vědci jako Robson (Robson, 1985), Peters (Peters, 1963) a Mausersberger (Mausersberger, 1954). Dospělý bourec morušový svinuje kokon a uvolňuje viskózní tekutinu produkovanou dvěma hedvábnými žlázami. Tato látka je vylučována párovými vývody, které jsou umístěny v hlavě bource morušového a plní funkci zvlákňovacího aparátu. Viskózní složka (fibroin) je obalena další vrstvou (sericin) vytékající z hedvábných žláz. Produktem takové spřádací činnosti se stává hedvábné vlákno – sloučenina sericinu a fibroinu. Sericin neboli „hedvábné lepidlo“ je menší ze dvou složek hedvábného vlákna (tvoří asi 25 % celkové hmoty surového hedvábí) a obsahuje řadu dalších prvků, jako jsou vosky, tuky a pigmenty.
Sericin je žlutá, křehká a nepružná látka, u které bylo prokázáno, že má baktericidní vlastnosti (Chang, 2005). Působí jako spojovací materiál pro dva prameny fibroinu a skrývá jeho jedinečnou schopnost odrážet světlo. Sericin je známý svou amorfní strukturou a lze jej od fibroinu oddělit procesem nazývaným „ošetření horkým roztokem“. Komatsu (Robson, 1985; Gulrjani, 1992) uvádí, že další zpracování sericinu mu umožní izolovat látky Sericin I, Sericin II, Sericin III a Sericin IV kvůli jejich různé rozpustnosti v horké vodě. Nejvíce sericinu je obsaženo ve vnějších vrstvách kokonu, a tedy čím blíže ke středu, tím méně.
Fibroin – hlavní prvek hedvábí, přesně ten, který jde do výroby. Jedná se o ve vodě nerozpustný protein (dosahuje 75 % celkové hmotnosti surového hedvábí). Textura fibroinu a jeho síla jsou dány jeho krystalickou strukturou, přes kterou si fibroin zachovává velké množství aminokyselin.
Kokony se nejprve pečlivě třídí podle odrůdy, velikosti, kvality a případných vad. Jedna z prvních operací při výrobě hedvábí – zakuklení (Das, 1992). Jedná se o technologický postup získávání surového hedvábí navíjením nití z kokonů. Pro navíjení kokonů se vytříděné kokony napařují, aby změkly sericinové nitě, které je slepují dohromady. Kukla bource morušového během této léčby zemře a je zlikvidována. Kokony se shromažďují za konce vláken vyčnívajících na hladině vody, rozmotávají je a proplétají dohromady do jediné hedvábné nitě (surové hedvábí) o hmotnosti od 20 do 25 denier (ekvivalent hmotnosti 9 km hedvábí v gramech). ). Celý tento proces se obvykle provádí na automatických navíjecích strojích.
Často se takto vzniklé nitě podrobují další úpravě: tzv. „kroucení“, tzn. vytvoření nové nitě stočením několika nití surového hedvábí do jedné (Dixit, 1990). Předené hedvábí se používá k výrobě speciálních tkanin, které jsou obvykle tkané nebo pletené.

Proces odlepování je klíčová při mokrém zpracování surového hedvábí, protože surové hedvábí se skládá ze dvou složek, fibroinu a sericinu, které ho obalují. Sericin obsahuje některé nečistoty jako vosk, tuk, minerální soli a pigmenty. Přestože sericin obsahuje stejné aminokyseliny jako fibroin, jejich podíly v obou látkách nejsou stejné. V tomto ohledu musí být postup degumování hedvábí prováděn s velkou opatrností, aby nedošlo k poškození fibroinových vláken. Odlepení je nutné:
- K oddělení sericinu od fibroinu. Fibroin by neměl být ovlivněn;
- Chcete-li se zbavit nečistot (vosk, tuk a minerální soli), které narušují proces barvení nebo tisku;
- Zvýšit adsorpční kapacitu vlákna vůči barvám a chemikáliím;
- Aby fibroin získal lesk (Saligram, 1993).
- Degumování vodou pod tlakem (115°C);
- Degumování mýdlovým roztokem (98°C);
- Degumování syntetickým detergentem (98°C);
- Degumování kyselinami;
- Degumování pomocí enzymů.
- Enzymy jsou komplexní proteiny s vyšší molekulovou hmotností;
- Enzymy jsou citlivé na teploty a kyselosti, které překračují jejich přípustné hodnoty, protože pod jejich vlivem mohou změnit své vlastnosti;
- Enzymy reagují se specifickými částmi substrátu v závislosti na aktivních místech a typech;
- Enzymatické reakce jsou reverzibilní.
- Gravimetrická metoda (Sandoz Color Chronicle, 1991; Shukla et al., 1992);
- Barvící metody, které používají různá barviva k detekci přítomnosti sericinu. Typicky se používají následující barviva: C.I Direct Red 2, C.I Direct Blue 22, C.I Direct Red 61, C.I Direct Green 9 (BianchiandColonna, 1992);
- Stupeň poškození vlákna lze posoudit podle viskozity výsledného roztoku, elasticity vlákna a složení jeho nejjednodušší aminoskupiny;
- Účinnost degumování lze posoudit pomocí rastrovacího elektronového mikroskopu (SEM).
Motýl cikánský se vyznačuje ostrým sexuálním dimorfismem. Samec má rozpětí křídel 35-40 mm, je tmavší barvy než samice, přední křídla jsou hnědošedá s příčnými tmavými vlnitými pruhy. Zadní křídla jsou jednotně hnědá. Břicho je úzké, tykadla jsou široce česaná. Samice je větší, s rozpětím křídel 55-70 mm. Oba páry křídel jsou špinavě bílé, na předních jsou výrazné tmavě hnědé klikaté pruhy (obr. 1). Tělo samice je velké, břicho nápadně ztluštělé a hustě pokryté hnědožlutými chlupy.

Rýže. 1. Dospělý cikánský můra (foto: John H. Ghent, USDA Forest Service, Bugwood.org)
Tykadla jsou setiformní, jednostranně česaná. Vajíčka jsou zpočátku narůžovělá, později tmavě šedá, kladená v zaoblených hromádkách, pokrytá světle žlutohnědými chloupky z těla samic (obr. 3). Tyto útvary obsahující 500-1000 vajíček se nazývají „houby“. Chloupky chrání vajíčka před mrazem (vajíčka přezimují) a také je činí neviditelnými v okolí (mimikry).

Rýže. 2. Housenka zavíječe (foto: Koltunova E.V.)

Rýže. 3. Ovipozice zavíječe na bříze v březových lesích
lesy Trans-Uralu (foto: Koltunova E.V.)
Housenky mají charakteristickou nažloutlou, tmavě mramorovanou kresbu, dosti velkou hlavu se dvěma hnědými pruhy. Tělo je válcovitého tvaru. Mladé, čerstvě vylíhlé housenky jsou černé a pokryté chlupy. Housenky prvního instaru mají na druhém, třetím a desátém segmentu těla nažloutlé skvrny. Dospělé housenky jsou dlouhé 40-80 mm. Základní barva je šedá až žlutohnědá se třemi tenkými žlutými linkami nebo jednou širší podélnou tmavě hnědou linkou na hřbetě. Prvních pět segmentů má 2 modrofialové bradavice, všechny ostatní mají 2 červené bradavice (obr. 2). Kukly jsou mumiformní, tmavě hnědé, matné, dlouhé 20-30 mm, s řídkými chomáčky chlupů a háčky na konci těla. Již na kuklích je možné určit budoucí pohlaví, kromě jiného, podle genitálních otvorů. Na rozdíl od bource morušového jsou chlupy na hřbetní straně hnědožluté.
Cikán poškozuje většinu listnatých stromů (více než 300 druhů). Housenky nejčastěji napadají ovocné stromy, ze kterých se snadno šíří na dub, habr, lípu a četné další druhy. Ohniska hromadného rozmnožování tohoto škůdce se nejčastěji vyskytují v jižních oblastech na dubových plantážích.
Motýli se objevují v přírodě od konce srpna do září a v teplých oblastech – o měsíc dříve. Samci začínají létat jako první, o několik dní dříve než samice. Samice jsou těžké, většinou nelétají a sedí na kůře kmenů. Samci, zejména večer, energicky poletují a hledají samice, které je přitahují, a z poměrně velké vzdálenosti sexuální atraktanty (feromony). Oplozené samice kladou vajíčka v hromadách na kůru spodní části listnatých stromů, za normálních podmínek do výšky maximálně 4 m. Jedna samice naklade průměrně 250-500 vajíček. Motýli žijí jen asi dva týdny. Přezimovaná vajíčka obsahují malé housenky, které vylézají až v dubnu následujícího roku, kdy průměrná teplota dosáhne 10 °C. Zpočátku se malé housenky neživí, žijí pospolu a teprve po pár dnech se plazí po koruně. Housenky prvního instaru ohlodávají malé otvory v mladých listech, zatímco starší housenky požírají listy zvláštním, pro ně charakteristickým způsobem. Často zbytky listů padají na zem. Když je nedostatek potravy, housenky jedí nezralé pletivo výhonků, květů a poupat. Bylo zjištěno, že housenky středního věku v porostech s podrostem a vlhkou lesní podestýlkou často migrují po stromech.
Mladé housenky se živí převážně ve dne, k intenzivnímu krmení starších housenek (od 4. a výše) dochází především v noci a pouze v případě propuknutí hromadné reprodukce – ve dne. Asi po 70-80 dnech, v teplých jižních oblastech po 45-50 dnech, po intenzivním krmení, jsou housenky připraveny k zakuklení. Kuklení nastává v polovině července, na jihu – již koncem června. Fáze kukly trvá přibližně 10-15 dní.
V řídkých lesích nalézá můra cikán optimální podmínky pro svůj vývoj. Tento druh je nebezpečným škůdcem zejména proto, že obvykle poškozuje nerovnoměrně širší plochy a dále se šíří v průběhu gradace, obvykle v období tří až čtyř let. Frekvence gradací ve středním pásmu je 5-8 let. Většina stromů se zotaví z poškození, které je nejintenzivnější na konci června a července, a znovu se zazelenají. Pokud bylo v kritickém období velmi sucho, odumírají jednotlivé duby, někdy i celé dubové porosty. Při ohniscích hromadného rozmnožování se můra cikánská vyskytuje nejen v korunách stromů, ale také v kudlích a mladých lesích.
Cikán je rozšířen v teplých oblastech po celé Evropě (kromě severních oblastí), hranice rozšíření jde přes Středomoří do severní Afriky. Známý je také v severním Mongolsku, Číně (až po hranici palearktické oblasti), dále v Koreji a Japonsku. V 19. století byl z Evropy zavlečen do Severní Ameriky (Massachusetts), kde našel vhodné podmínky pro svůj rozvoj a až do roku 1952 se rozšířil téměř po celých Spojených státech a stal se zde nebezpečným škůdcem.
V Rusku je distribuován po celé evropské části. Severní hranice probíhá od linie Petrohrad – Volgograd – Perm. Vyskytuje se na Krymu a Kavkaze, Uralu, Altaji, severním Kazachstánu, Střední Asii, Sibiři a Dálném východě.
Začátkem roku 2007 operovala ohniska můry cikánské na ploše 813,2 tisíc hektarů ve 30 regionech Ruska. Hlavní osídlené oblasti se nacházely v oblasti Dálného východu (600,5 tisíc hektarů). Tam byla největší ohniska škůdce, která působila na území Primorsky (490,0 tis. hektarů) a Chabarovsku (110,2 tis. hektarů). a Čeljabinské oblasti.
V lesích regionu Sverdlovsk je fytofág pravidelně pozorován, ohniska se vyskytují v oblastech nepřesahujících 3,5 tisíce hektarů. V autonomním okruhu Chanty-Mansijsk a Jamalo-Něnec nebylo po celou dobu pozorování pozorováno žádné ohnisko škůdce. Maximální plocha osídlená cikánskými můry byla identifikována v roce 1996 na plantážích v oblastech Kurgan (165,0 tis. hektarů) a Ťumeň (260,9 tis. hektarů). Hlavními poškozenými druhy v regionu jsou bříza a dub letní.
Dynamika propuknutí masové reprodukce cikánů
bource morušového v Trans-Uralu
Charakteristiky populační dynamiky můry v lesích Zauralské oblasti jsou studovány od 50. let 20. století (Raspopov, 1961, 1970, 1977; Raspopov, Rafes, 1978; Sokolov, 1988, 1990; Koltunov; a kol., 1992, 1998; Koltunov, 1993, 1996, 2000, 2001).
V lesích Zauralské oblasti bylo registrováno 5 po sobě jdoucích ohnisek hromadného rozmnožování zavíječe v březových lesích. V Čeljabinské oblasti došlo k prvnímu ohnisku masové reprodukce v letech 1953-1958, poté v letech 1963-1969, 1973-1980, 1984-1990 a malá ohniska se objevila v roce 2003 v lesích sousedících s Baškortostánem, kde došlo k intenzivnímu propuknutí masové reprodukce. rozvinutý. Ve zbytku Čeljabinské oblasti nebyla žádná ohniska.
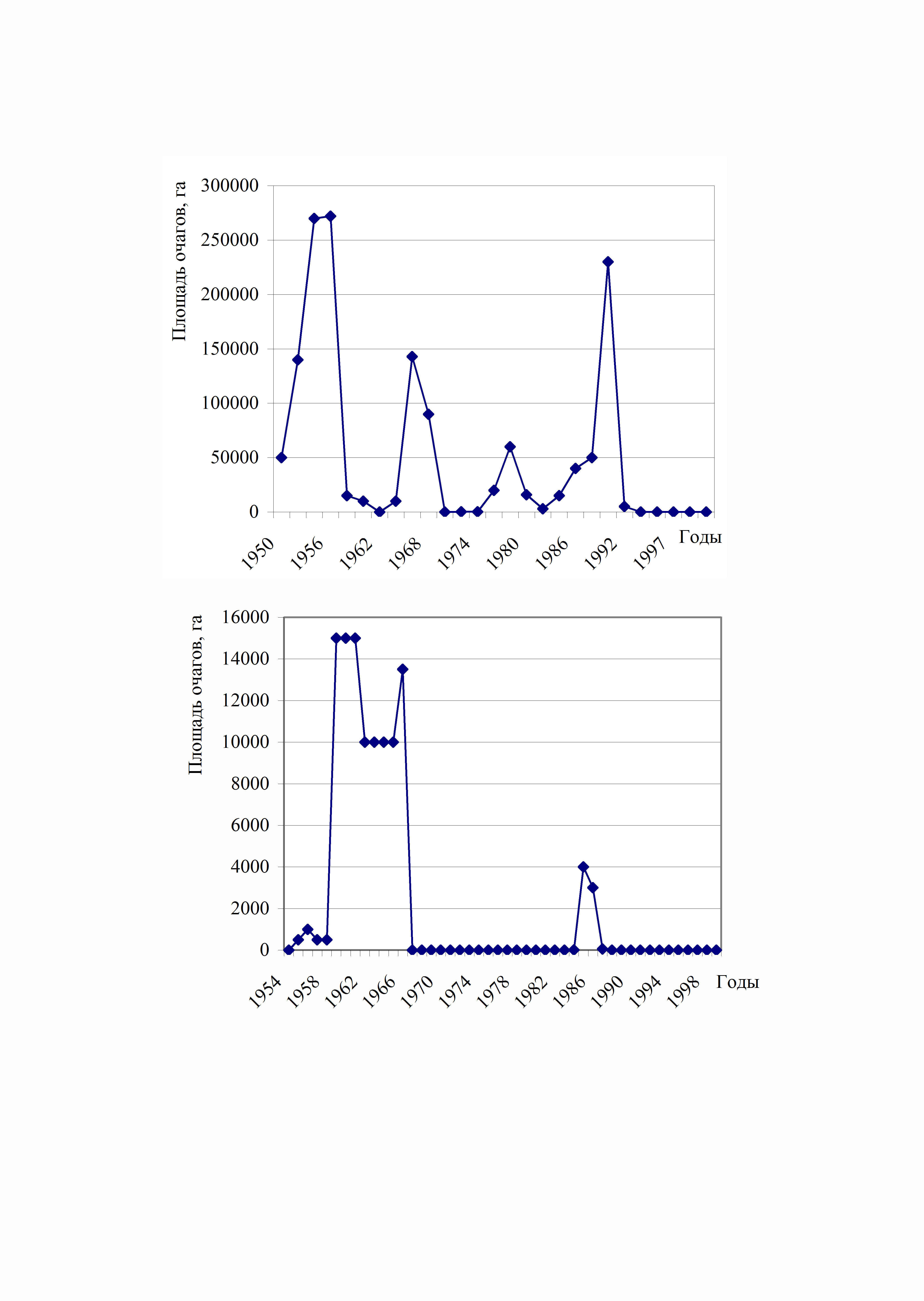
Rýže. 4. Dynamika propuknutí masové reprodukce cikánů
bource morušového v Čeljabinsku (horní) a
Sverdlovské (dolní) oblasti
Největší a nejdelší ohnisko masové reprodukce bylo v letech 1953-1958. To je způsobeno synchronizací ohniska se zvláště intenzivními a dlouhotrvajícími jarními a letními suchy. V důsledku dlouhodobého působení abiotického stresového faktoru na porosty stromů byl pozorován i částečný úbytek březových lesů (Raspopov, 1961, 1970, 1973; Koltunov, 1993,1996, 1992; Koltunov et al., 1998, 1984). Druhým nejintenzivnějším ohniskem bylo vypuknutí masové reprodukce v letech 1992-1999. Propuknutí bylo také synchronizováno se suchem (Koltunov, Andreeva, 5). Vztah mezi propuknutím masové reprodukce a suchem je podrobněji rozebrán v kapitole 9. Četnost propuknutí masové reprodukce zavíječe v lesích Čeljabinské oblasti byla 11-1992,1998 let (Koltunov et al., 1993; Koltunov, XNUMX). Preferované druhy: bříza, osika.
V lesích regionu Sverdlovsk došlo k prvnímu ohnisku hromadné reprodukce cikánského můry v roce 1954 a trvalo až do roku 1970. (obr. 4). Další ohnisko nastalo až v roce 1986 a vymřelo v roce 1990. Populace zavíječe žije pouze v jižní a jihozápadní části regionu (Koltunov, 1993). Je to dáno příznivějšími klimatickými podmínkami v této zóně. Frekvence je v průměru 20 let. Je také pozoruhodné, že celkové plochy ohnisek nejsou srovnatelné s ohnisky v Čeljabinské oblasti, protože jsou mnohonásobně nižší.
Obecně platí, že stejná asynchronie je pozorována ve skupině transuralských center hromadného rozmnožování zavíječe (obr. 5). Pokud tedy v roce 1990 došlo v Čeljabinské oblasti k velkému masovému rozmnožování můry cikánské. již vymírá, v lesích Ťumeňské oblasti se teprve v tomto období začíná rozvíjet (obr. 5). Stejný vzorec je pozorován v lesích oblasti Kurgan, pouze zde je zvláště dobře vyjádřena erupční fáze.
Na začátku roku 2007 v lesích regionu Kurgan. Bylo identifikováno 21.275 64.382 ha ohnisek hromadné reprodukce zavíječe, ke konci roku – 115 223 hektarů; v lesích Sverdlovské oblasti. na začátku roku bylo identifikováno 19105 hektarů ohnisek, na konci – 94537 hektarů; v lesích Ťumeňské oblasti. na začátku roku bylo objeveno 2007 15404 hektarů ohnisek zavíječe, na konci roku – 23196 2007 hektarů; v lesích Čeljabinské oblasti. na začátku roku XNUMX bylo identifikováno XNUMX XNUMX hektarů ohnisek, na konci roku XNUMX XNUMX hektarů (Rosleskhoz, XNUMX).
Pro studium faktorů asynchronie jsme studovali hlavní abiotické faktory ve fázi deprese, začátku rozvoje ohniska, v eruptivní fázi a fázi útlumu na příkladu ohnisek masové reprodukce zavíječe v Čeljabinské oblasti (Koltunov a kol., 1998; Koltunov, 2006). Přesvědčivě se ukázalo, že charakter vývoje ohniska je dán typem a sledem kombinace abiotických faktorů, které se výrazně liší nejen v různých regionech, ale i v podmínkách sousedních meteorologických stanic ve stejném regionu.
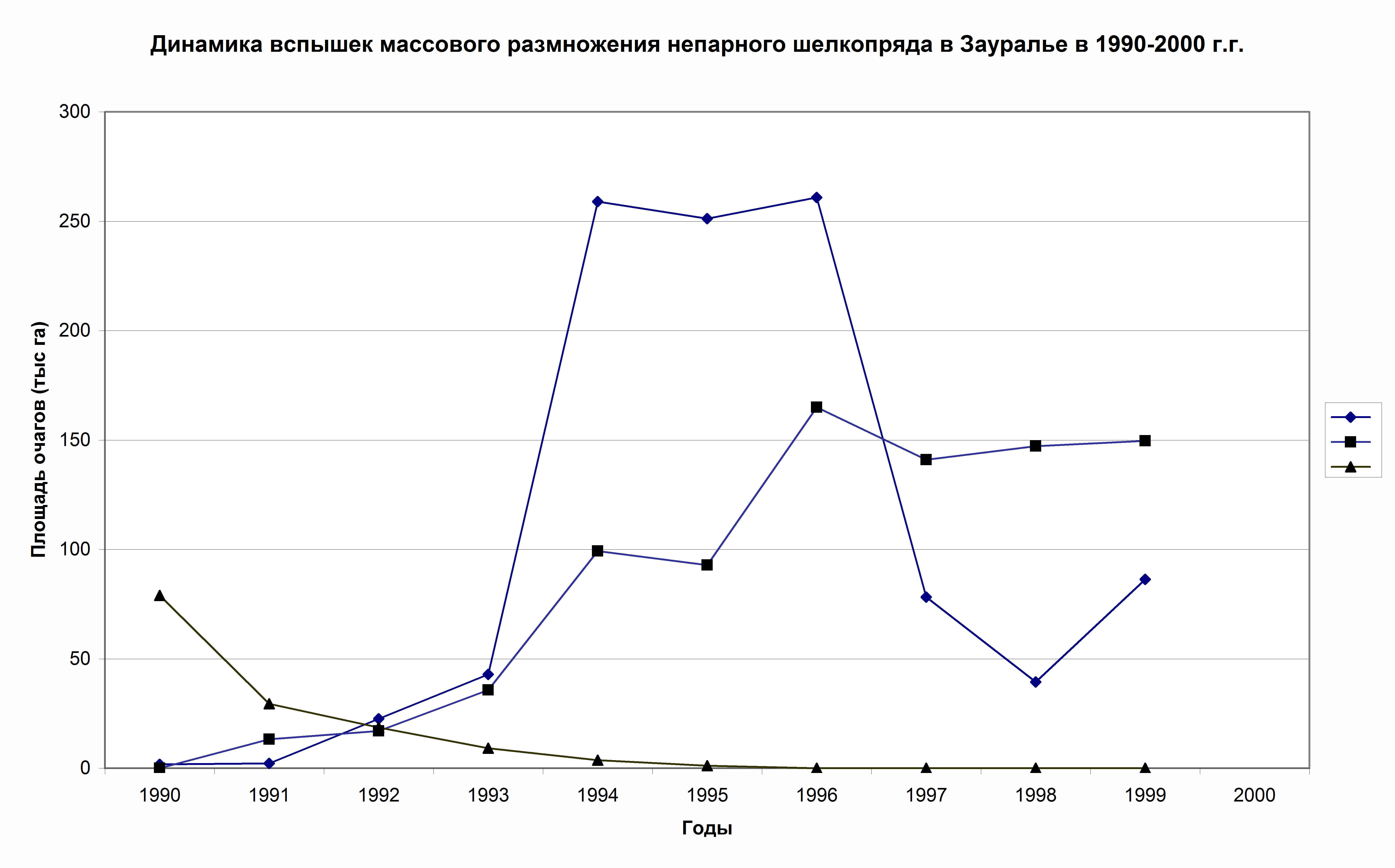
Rýže. 5. Charakteristiky populační dynamiky zauralských (▲), baškirských (■) a Ťumeňských (♦) geografických populací můry jíkovité v letech 1990 až 2000 (Koltunov, 2006)
Místní prostorová struktura ohnisek je tedy dána zvláštnostmi místního projevu klimatických podmínek (jarní-letní sucha a horké větry, míra jejich intenzity, trvání), které se vždy liší i v podmínkách dvou sousedních regionů. . Čím intenzivnější a déle trvající sucha, tím vyšší je potenciál pro ohniska a oblast ohnisek a tím menší jsou rozdíly v ohniscích v přilehlých regionech. Poměrně přesvědčivým příkladem je vypuknutí hromadného rozmnožování můry jíkovité v Zauralu v 50. letech, kdy byla pozorována zvláště intenzivní, stabilní a dlouhotrvající sucha (Koltunov, 1993, 1996, 2006; Koltunov et al., 1998). Tento problém je podrobněji rozebrán v této monografii v kapitole 5.
Asijská forma můry cikánské chybí nejen v evropské části Ruské federace, ale ani v Evropě a Severní Americe. V zemích tichomořské oblasti je tento druh v karanténě. V těchto zemích je zařazení můry na seznam karanténních druhů naprosto oprávněné, protože se v zemích buď nevyskytuje, nebo má omezené rozšíření.
V roce 1992 byla asijská forma můry cikán zařazena do „Seznamu rostlinných škůdců, rostlinných patogenů, rostlin (plevelů) karanténního významu pro Ruskou federaci“ jako předmět vnější karantény. V současné době probíhají pokusy o zavedení vnitřní karantény na územích Primorsky a Chabarovsk, jakož i v Republice Sakha a některých zakládajících entitách okresu Volha kvůli výskytu ohnisek můry cikánských v těchto oblastech. Podle našeho názoru, který se opírá o výsledky dlouholetého výzkumu ekologie strakáče (Koltunov, 1993, 1996, 2006; Koltunov et al., 1998), zařazení této formy do seznamu karanténních objektů byla zcela neopodstatněná. Pro specialisty, kteří by rádi získali o této problematice úplnější informace, odkazujeme na naši publikaci (Koltunov E.V., Ponomarev V.I., Fedorenko S.I. O zavedení karantény proti asijské rase cikánského můry // Lesnictví, č. 4, pp 43-46, 2001), ve kterém je tato problematika velmi podrobně rozebrána. Článek také uvádí, dle našeho názoru, celkem přesvědčivé argumenty o mylnosti tohoto rozhodnutí. Rosleskhoz v současnosti zastává podobnou pozici (Rosleskhoz, 2007). Bohužel je třeba poznamenat, že zavlečení této formy můry do karanténních zařízení způsobuje Rusku poměrně značné ekonomické škody, protože jsou vyžadována poměrně nákladná karanténní opatření.
Doporučení pro ochranu lesů před cikánkou
Není pochyb o tom, že odborníci na ochranu lesa zajímající se o odbornou literaturu k tomuto objektu dobře znají náš názor na současnou podivnou situaci s bojem proti ohniskům zavíječe v březových lesích (Koltunov, 1993, 1996; 2006; Koltunov, Fedorenko, 1998; Koltunov a kol., 2006). Jak známo, bříza má v příznivých lesních podmínkách mimořádně vysokou úroveň entomotolerance. Proto při jediné úplné defoliaci procento vyschnutí obvykle nepřesáhne 2-5 % (Koltunov, 1993, 1996, 2006; Koltunov et al., 1998). To platí zejména pro odolnost vůči defoliaci fytofágním hmyzem skupiny jaro-léto. Výraznější lokální prosychání se nám podařilo zaznamenat pouze v březových lesích, které byly výrazně postiženy bakteriální vodnatelností po úplné defoliaci korun břízy zavíječem. Zvýšená úmrtnost po jednorázovém úplném odlistění břízy byla také pozorována lokálně a v důsledku nedostatku kyslíku v kořenovém systému břízy, vystavené úplné defoliaci na mikrodepresích reliéfu, v důsledku vydatných srážek v druhé polovině léta na půdách s jílovitými čočkami v březových lesích Zauralu (Koltunov, 1993, 1996). Ale tohle bylo velmi místní povahy. Obecně lze říci, že drtivé procento březových lesů v průběhu více než 20 let pozorování velmi úspěšně přestálo několik úplných defoliace korun a v současné době mají poměrně vysokou hustotu (obr. 6).

Rýže. 6. Úsek březového lesa v Kamensko-uralské oblasti, který v letech 1966, 1986 utrpěl silnou defoliaci zavíječem, letní a podzimní komplex fytofágů v roce 2000 a průměrnou úroveň defoliace v roce 2006, stejně jako slabou defoliaci. v letech 2007-2009
Zároveň byl lokálně zaznamenán úbytek výsadby v důsledku poškození zavíječem v roce 2007 na 53 hektarech v lesích Kurganské a Čeljabinské oblasti, kde byly výsadby již dříve oslabeny vlivem nepříznivých povětrnostních podmínek. . Celková plocha ohnisek však byla 64 tisíc hektarů (Rosleskhoz, 2007). Na územích Primorsky a Khabarovsk nebyly po celou dobu pozorování (od 50. let minulého století) zaznamenány žádné případy úmrtí výsadeb.
Hromadné ošetřování ohnisek proti zavíječi v březových lesích, které se v současné době provádí, je proto zcela ekologicky a vědecky neopodstatněné. V tomto ohledu lze pouze doporučit odmítnutí hromadného ošetření březových lesů v centrech hromadné reprodukce cikánských můr na území Uralského federálního okruhu.
Toto doporučení přitom vůbec nevylučuje, že za určitých podmínek na místní úrovni lze tato ošetření provádět, např. je-li nutné zabránit defoliaci březových lesů v okolí dětských táborů, domovů důchodců , sanatoria atd. objekty výhradně pro ochranu lesních ploch pro estetické a rekreační účely. Totéž platí pro městské lesoparky a veřejné rekreační oblasti. Prvky námi navržené nové strategie ochrany lesa (Koltunov, 1998, 2006) jsou uvedeny v samostatné části stránky.
Zcela odlišná ekologická situace se vyvíjí v oblastech masového rozmnožování zavíječe v dubových lesích. A přestože odborná literatura poskytuje zcela odlišné informace o úrovni entomotolerance dubu k defoliaci, složitá situace s dubovým biotopem v Rusku, zejména v jeho východní části, nesprávným lesním hospodařením v dubových lesích (mrostové hospodářství), intenzivní působení antropogenních faktorů a extrémně reálné nebezpečí silného dočasného oslabení dubu po silné defoliaci, které je často doprovázeno jeho poškozením infekčními chorobami, podle našeho názoru vyvolává vědecké diskuse na téma, zda je nutné léčit ohniska hromadného rozmnožování v dubových lesích irelevantní. Domníváme se, že na rozdíl od břízy je v dubových lesích v případě nebezpečí silného jednoduchého odlistění, nebo středního dvojitého odlistění vhodné ložiska ošetřit, aby se předešlo silnému dočasnému oslabení dubu a jeho částečnému vysychání a rozpadu. Stručnost doporučení v této části je způsobena téměř úplnou nepřítomností dubu v lesích Uralského federálního okruhu (až na velmi malé výjimky).















